Project 4
Post-transcriptional regulation of gene expression during neuronal development
Nascent mRNAs are co-transcriptionally assembled into mRNPs. In neurons, a subset of these transcripts is transported to peripheral cellular locations such as synapses for translation. Here, many heterogeneous nuclear ribonucleoproteins (hnRNPs) play an important role as they interact co-transcriptionally with the RNA through specific binding sites or cis-acting elements located in the untranslated regions (UTRs). These interactions contribute to sort transcripts for cytoplasmic transport and localization. This post-transcriptional level of gene expression regulation is important during development and differentiation. Recently, the interest in this research area has been amplified by the discovery that newly synthesized RNA transcripts are methylated, a modification that seems to have a general impact on gene expression during neuronal development and differentiation. A set of methyl transferases and demethylases regulate the levels of such modifications.
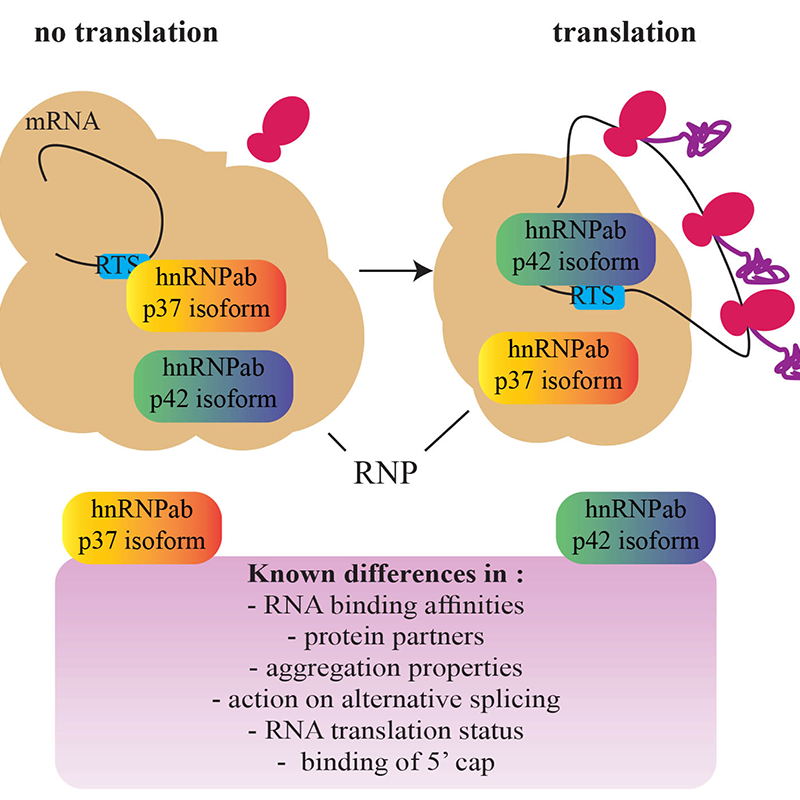
An ongoing project in the lab focuses on how RNA transcripts are regulated by methylation at transcriptional hubs. We work on the hypothesis that RNA methylation is important to regulate the cytoplasmic fate of transcripts. We study a potential reader of RNA methylation, the hnRNP protein CBF-A/hnrnpab. This protein interacts with actin within the nascent pre-ribonucleoprotein complex and it is important for cytoplasmic RNA trafficking in oligodendrocytes, hippocampal neurons, and spermatogenic cells. Selective binding of the two CBF-A/hnrnpab isoforms p37 and p42 to the RNA trafficking sequence (RTS) in the RNA 3’ untranslated region regulates translatability during development. By binding the RTS, p37 and p42 respectively stabilize translationally repressed mRNA for transport and translationally active mRNA for localized translation (Figure 3).

We currently study if the two isoforms are readers of RNA methylation, and if this mechanism contributes to neuronal development by regulating cytoplasmic mRNA trafficking. To address this question we apply genome-wide approaches such as RIP-seq and global transcriptional profiling of embryonic fibroblasts from a mouse model where both CBF-A/hnrnpab alleles are disrupted. To find out the impact of these mechanisms on development we combine the analysis of primary neurons from the knockout mouse model with reprogramming of embryonic fibroblasts from the CBF-A/hnrnpab knockout mouse. We study the role of RNA methylation on the CBF-A/hnrnpab binding affinities in embryonic fibroblasts exhibiting RNA hypermethylation.
A long term goal is to correlate the above mechanisms with compromised learning and memory in vivo using the available mouse model.