-
Academics
-
Admissions
 Your journey to NYUAD starts here. Attend an application workshop or information session.Admissions Events
Your journey to NYUAD starts here. Attend an application workshop or information session.Admissions Events -
Research
-
Campus Life
 Live the possibilities. Be part of a dynamic community of students from over 115 countries.Take a Tour
Live the possibilities. Be part of a dynamic community of students from over 115 countries.Take a Tour - Public Programs
- About
- News
- Events
- Social Media Directory
- Press Room
- Faculty
- Current Students
- Alumni
- عربي


Project 1
Actin and myosin in genome organization and transcriptional reprogramming during development
Increasing evidence suggests an important role for nuclear actin in heterochromatin segregation and maintenance. We recently reported that β-actin regulates heterochromatin by controlling genomic deposition of Brg1, the ATPase subunit of the chromatin remodeling complex BAF (Figure 1). We found that his has a major impact on the way the genome is organized, on global transcription and cell identity.
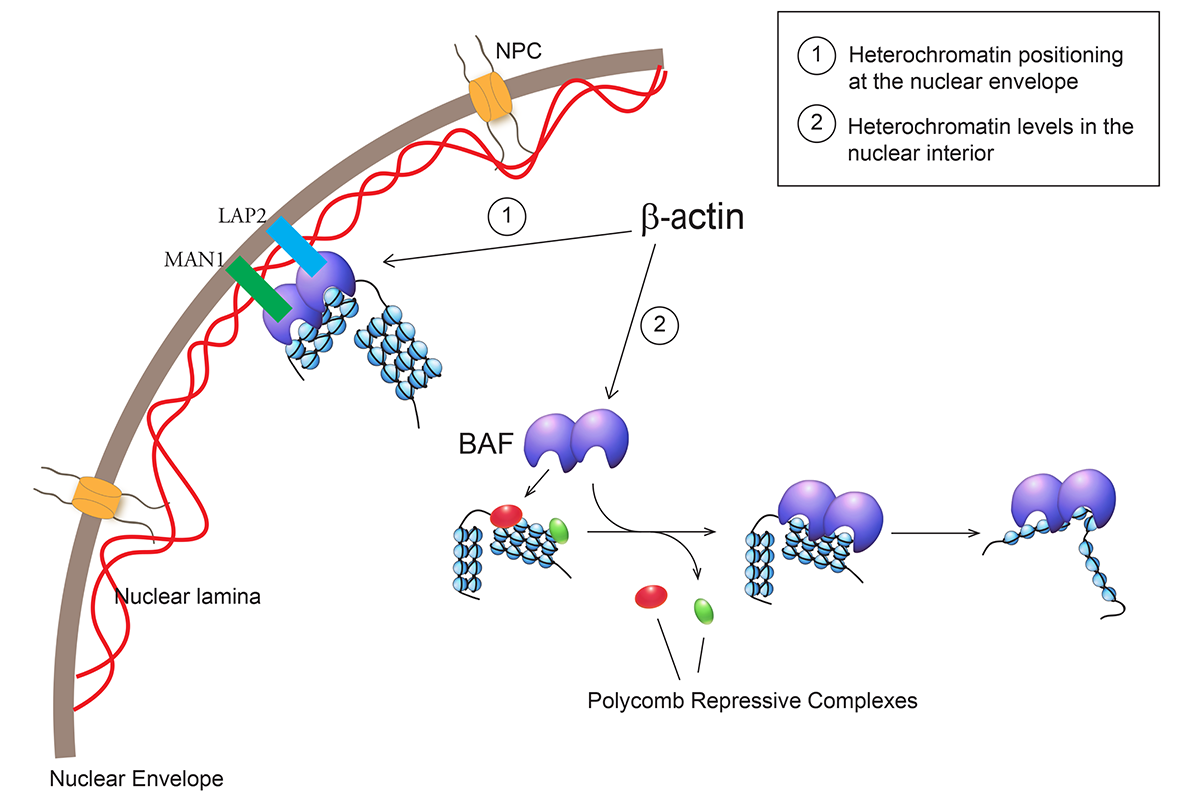
.")
Regulation of heterochromatin is known to be important during development and differentiation. In particular, the extent of histone H3 methylation on lysine 9 (H3K9me3) is considered as barrier during development and differentiation. In light of this, we have started to address how actin-dependent heterochromatin regulation controls H3K9me3 levels and affects transcriptional reprogramming during neuronal development. Interestingly, we found that the absence of β-actin does not compromise transdifferentiation of embryonic fibroblasts to neurons. However, the altered heterochromatin reorganization and transcriptional reprogramming leads to neuronal subtype switches. These results suggest that nuclear actin has an important role in presetting the chromatin during neuronal development, facilitating activation of occluded genes.
Ongoing projects are geared towards studying the role of nuclear actin in the topological organization of chromatin and how this is implicated in neuronal development. We address these questions using a combination of genome-wide approaches (Hi-C and ATAC-seq) and reprogramming of fibroblasts with compromised β-actin expression to neuronal cells. We are also working on other transdifferentiation models to study adipogenesis and osteogenesis and methods to dedifferentiate fibroblasts to induced pluripotent stem cells, to generalize on the importance of actin-dependent heterochromatin regulation.
Similar projects are ongoing to address the role of a specific myosin 1c isoform - nuclear myosin 1 (NM1) - in genome organization. We and others reported the presence of a nuclear actomyosin complex localizing to active transcription sites. We discovered that NM1 is required to engage the polymerase in elongation of nascent RNAs. Further, we showed that NM1 is a component of the multiprotein assembly B-WICH, containing the chromatin remodeling complex WICH with the subunits WSTF and the ATPase SNF2h. NM1 binds to the chromatin via its C-terminal tail domain and appears to act as a molecular switch interacting either with polymerase-associated actin or SNF2h. This molecular switch activates both RNA polymerase I and RNA polymerase II transcription. Given that NM1 is enriched at transcription start site of a large number of genes, we have suggested that NM1 establishes a chromatin landscape for transcription activation.
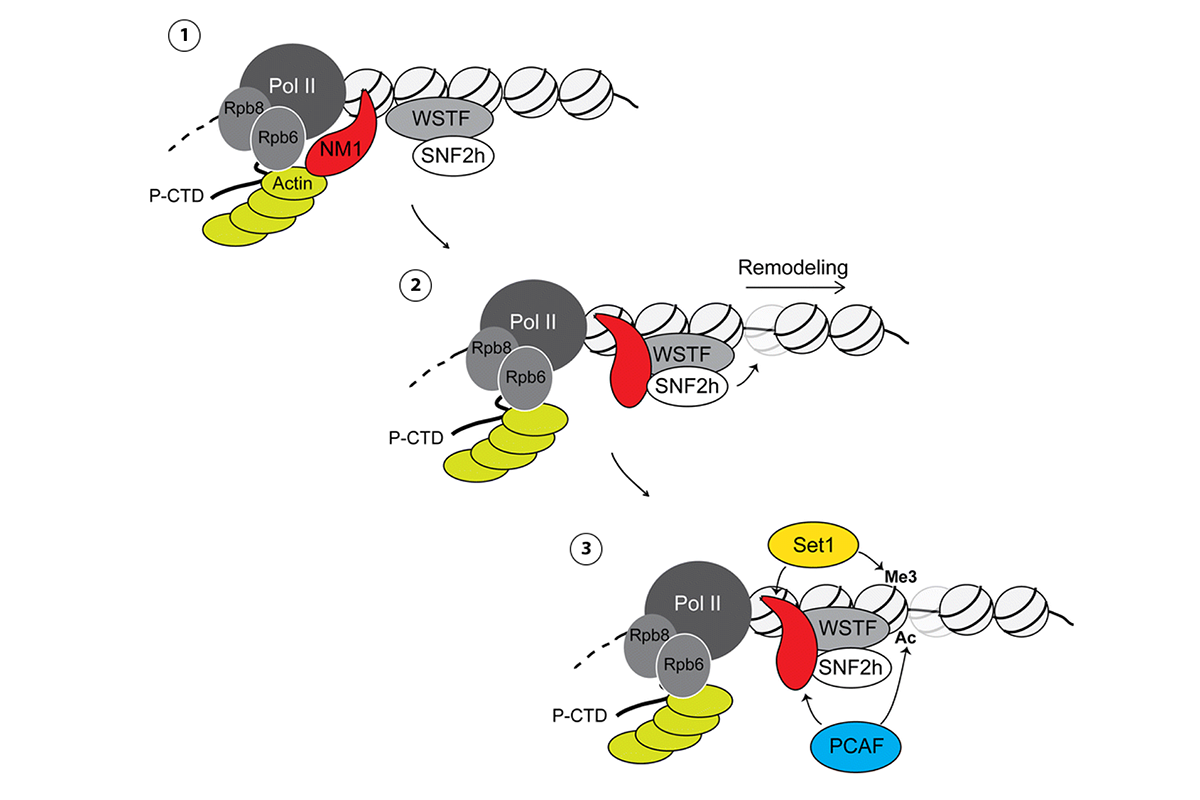
The above studies have not yet established whether myosin-dependent regulation occurs at the gene level only or also contributes to the 3D organization of the genome, for instance by facilitating establishment of transcriptional hubs. To address these questions, we are currently working with a knockout model for NM1. Using this system, ongoing projects aim at performing genome-wide analyses (primarily based on Hi-C and ATAC-seq) in combination with transcriptional profiling of NM1 knockout embryonic fibroblasts. To find out if these NM1-dependent mechanisms are important during development and differentiation, part of this project focuses on reprogramming embryonic fibroblasts with compromised NM1 expression to neuronal cells.